

Tipos de Sinalização
Sinalização Parácrina
Nesse tipo de sinalização, as moléculas sinalizadoras extracelulares produzidas pela célula sinalizadora difundem-se localmente, interagindo apenas com células-alvo próximas à célula sinalizadora. Essas moléculas têm difusão restrita e são eliminadas rapidamente por enzimas, ou são retidas pela matriz extracelular

Ex: A sinalização parácrina acontece, por exemplo, na atuação do Óxido Nítrico (NO) na dilatação dos vasos sanguíneos. A acetilcolina liberada do terminal nervoso da parede do vaso, atua na célula endotelial e estimula a formação do NO a partir da arginina. O NO difunde-se rapidamente pela membrana da célula sinalizadora e se liga à proteína-alvo intracelular na célula muscular lisa da parede do vaso sanguíneo. Essa interação induz o relaxamento rápido da célula muscular lisa.
Sinalização Autócrina
A sinalização autócrina é um tipo de comunicação parácrina. Nesse tipo específico de sinalização, as células podem também responder às moléculas sinalizadoras que elas mesmas produzem.

EX: A sinalização autócrina leva à estimulação da proliferação e sobrevivência celular no câncer.
Esse tipo de sinalização também é importante durante o desenvolvimento embrionário. Por exemplo, quando uma célula decide-se por uma determinada via de diferenciação, os sinais autócrinos reforçam a decisão celular.
Sinalização Telécrina (ou endócrina clássica)
Nesse tipo de sinalização celular, as moléculas sinalizadoras liberadas pela célula sinalizadora são chamadas de hormônios. As células sinalizadoras, por sua vez, são chamadas de células endócrinas. Os hormônios produzidos pelas células endócrinas podem ser distribuídos por todo o corpo, já que são secretados na corrente sanguínea. As células-alvo para esses hormônios apresentam o receptor específico.
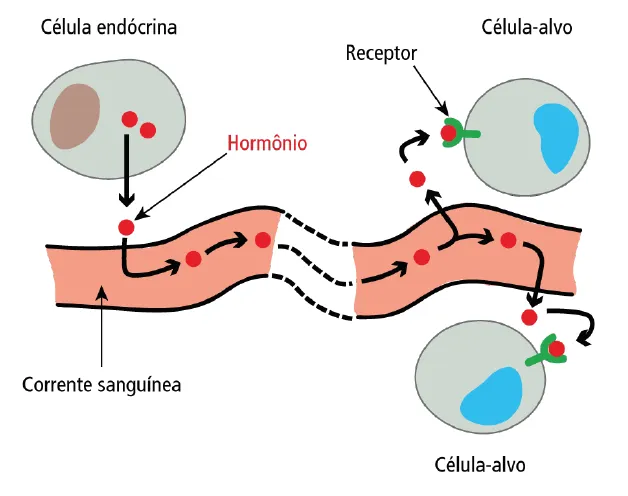
EX: Quando ocorre o aumento da glicemia sanguínea, por exemplo, as células β (célula endócrina) do pâncreas secretam insulina (hormônio). Em alguns minutos, a insulina é distribuída pela corrente sanguínea e atua em células-alvo estimulando a captação de glicose sanguínea. Com isso, há uma rápida redução da glicemia.
Sinalização Neurócrina
Através dos axônios, os neurônios estabelecem contato com células-alvo distantes. As moléculas sinalizadoras secretadas pelos neurônios são chamadas neurotransmissores. Os neurotransmissores são liberados de forma rápida e atuam de modo específico em uma célula-alvo.

EX:Quando um neurônio motor é ativado ele envia sinais elétricos e, ao chegar no terminal do axônio, esses sinais são convertidos em um sinal químico (neurotransmissor). A liberação da acetilcolina na junção neuromuscular, por exemplo, atua em receptores da fibra muscular (célula-alvo) estimulando a liberação de Cálcio (Ca2+) do retículo sarcoplasmático para o citosol. O Ca2+ associa-se com proteínas contráteis e, desse modo, estimula a contração muscular.
Sinalização Dependente de contato
Esse tipo de sinalização acontece, por exemplo, no período embrionário durante a diferenciação neural. Quando uma célula ectodérmica inicia DEPENDENTE DE CONTATO seu processo de diferenciação em célula neural, ela irá inibir as células vizinhas a seguir o mesmo caminho de diferenciação. Isso acontece pela interação do ligante Delta, que aparece na superfície da célula que está passando pelo processo de diferenciação neural, com a proteína receptora Notch das células vizinhas. Desse modo, a célula epitelial vizinha é inibida e não sofre a diferenciação neural.

EX: desenvolvimento neural

Resposta da célula
Uma célula produz um conjunto de receptores específicos e, por isso, mesmo com a exposição a inúmeras de moléculas sinalizadoras no microambiente celular, irá responder de forma seletiva. Ainda que a célula possua um determinado receptor, a resposta a uma determinada molécula sinalizadora não será a mesma entre diferentes células. Isso acontece porque o sistema de propagação intracelular e as proteínas efetoras são variáveis. A acetilcolina, por exemplo, induz diferentes respostas em tipos celulares especializados: Na célula marca-passo cardíaca reduz a velocidade de contração; na célula da glandular salivar, estimula a secreção; e na célula muscular esquelética atua estimulando a contração muscular. A célula responde simultaneamente a inúmeras moléculas sinalizadoras. Desse modo, combinações diferentes de moléculas sinais resultam em diferentes respostas. Na ausência desses sinais, a célula sofre apoptose.


A resposta a um sinal pode ser rápida ou lenta. O tempo de resposta é dependente do que acontece depois que a mensagem é recebida. Quando a molécula sinalizadora atua sobre uma via de sinalização celular que afeta a atividade de uma proteína já presente na célula alvo, a resposta será rápida. Entretanto, quando a resposta requer mudanças na expressão gênica, será lenta.
Mecanismo de Ação Hormonal
Todos os hormônios atuam através de receptores específicos presentes nas células-alvo. Os receptores fornecem o meio pelo qual os hormônios interagem inicialmente com as células, e podem se localizar na membrana plasmática, citosol e no núcleo celular. São proteínas as quais, os hormônios correspondentes, com alta especificidade e afinidade, provocam mudanças conformacionais que desencadeiam reações modificadoras do metabolismo da célula-alvo, constituindo a resposta celular. Os receptores não são componentes fixos, podendo variar o número de receptores para cada tipo de célula, com isso variando o grau de resposta. A união hormônio-receptor é forte, mas não covalente, sendo equivalente à união de um efetor alostérico com a enzima que o regula. O sítio de união é esteroespecífico, onde somente se une o hormônio correspondente ou moléculas similares. As estruturas análogas, denominadas “agonistas”, se unem aos receptores, ocasionando os mesmos efeitos que o hormônio. Em oposição aquelas estruturas, cuja união ao receptor não causa efeito hormonal, por bloquear os receptores, são chamados de “antagonistas”.
OBS: Existem dois tipos de mecanismos de ação hormonal
A) Os hormônios que possuem seus receptores na superfície externa da membrana plasmática das células-alvo, costumam exercer seus efeitos, pela alteração da permeabilidade da membrana, ou pela ativação de enzimas, a adenilciclase e a guanilciclase produzindo AMPc e GMPc respectivamente, conhecidos como “segundos mensageiros” e que têm suas concentrações aumentadas no interior da célula em resposta ao hormônio primário, regulando e modificando a velocidade de transcrição de genes específicos. Os hormônios deste grupo são transportados de forma livre pela corrente circulatória, sendo um mecanismo de ação mais rápido causando rápidas modificações metabólicas. O tempo de ação destes hormônios é de minutos ou segundos. As catecolaminas e os hormônios peptídeos utilizam este método.
OBS: A maior parte das moléculas sinalizadoras extracelulares são grandes e hidrofílicas e, portanto, atuam sobre receptores de superfície celular. Existem três grandes famílias de receptores de superfície celular: receptores acoplados a canais iônicos, receptores acoplados à proteína G e receptores acoplados a enzimas.
Nesse caso, o sinal será transmitido para dentro da célula de uma forma nova por meio de moléculas de sinalização intracelular. Essas moléculas intracelulares irão transmitir o sinal em cascata até a ativação de uma proteína efetora (Ex: enzima metabólica). Por outro lado, moléculas pequenas e hidrofóbicas atravessam livremente a membrana plasmática e poderão, portanto, atuar diretamente sobre receptores intracelulares.
OBS: As moléculas de sinalização intracelulares são ativadas em resposta à ligação de uma molécula de sinalização celular em um receptor presente na superfície celular. As vias de sinalização intracelular são responsáveis por transmitir e propagar o sinal para dentro da célula; amplificar o sinal recebido e torná-lo mais forte; integrar sinais de diferentes vias de sinalização; e distribuir o sinal para mais de uma via de sinalização ou proteína efetora.
A)
1- METABOTRÓPICOS
-
Receptores associados a proteína G
-
Receptores associados a enzimas
2- IONOTRÓPICOS (associados a canais iônicos)
Metabotrópico
1.1 -Receptores Associados a proteína G
OBS: A proteína G pode ativar enzimas e regular canais iônicos.
Muitas moléculas-sinal extracelulares atuam por meio de receptores associados à proteína G. Existem mais de 700 receptores associados à proteína G nos humanos. Esses receptores são formados por uma única cadeia polipeptídica transmembrana. A cadeia polipeptídica do receptor associado à proteína G atravessa a membrana plasmática sete vezes e há um domínio extracelular, onde a molécula sinalizadora extracelular se liga.

A proteína G, por sua vez, encontra-se na face interna da membrana plasmática. Ela é formada por três subunidades, alfa (α), beta (β) e gama(γ). As subunidades alfa e gama estão ligadas à membrana plasmática por meio de caudas lipídicas.
A interação do ligante com o domínio extracelular do receptor associado à proteína G causa uma mudança conformacional no receptor que permite que ele ative a proteína G. Antes da interação do ligante com o receptor, a subunidade alfa está ligada a uma molécula de GDP e encontra-se inativa. A mudança conformacional do receptor leva à ativação pela perda de afinidade da subunidade alfa por GDP e a troca por GTP
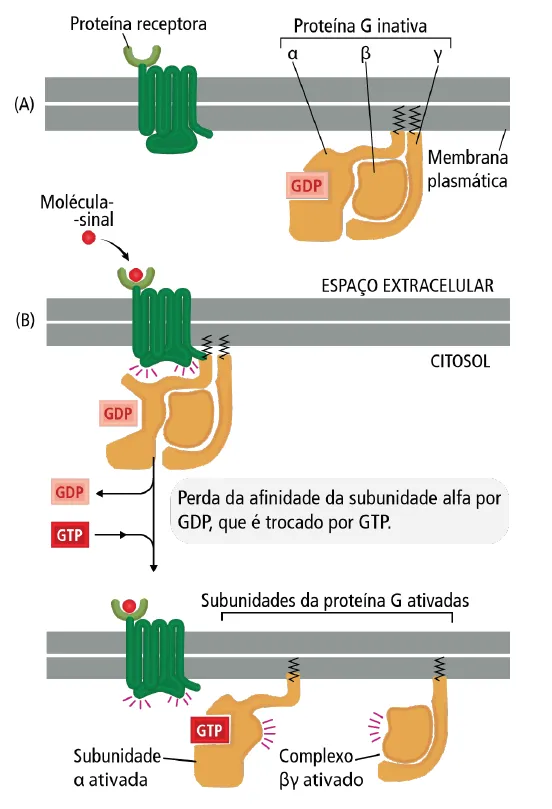
troca por GTP, ativa a subunidade α que, então, se dissocia do complexo βγ. Ambos podem, então, interagir com outras proteína-alvo na membrana plasmática (Ex: canais iônicos ou enzimas ligadas à membrana plasmática) para transmitir o sinal recebido. A subunidade alfa, que possui atividade GTPase, hidrolisa o seu GTP a GDP quando o sinal cessa.
Então, as subunidades se associam novamente.
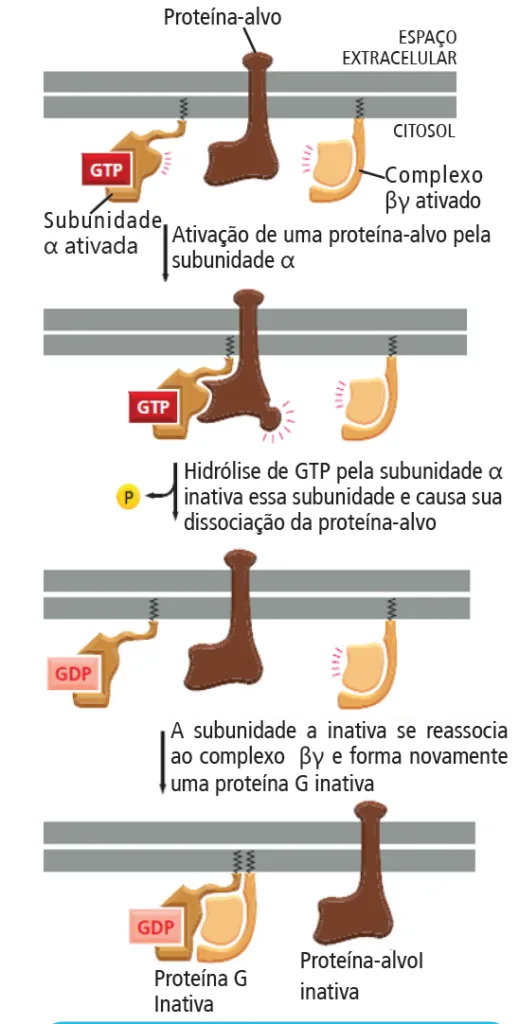
Um exemplo, é a proteína G acoplada à abertura de canais de K+ na membrana da célula muscular cardíaca. Quando a acetilcolina interage com um receptor associado à proteína G, ocorre a ativação da proteína G. A subunidade α se dissocia do complexo βγ, o qual se liga ao canal de K+ na membrana plasmática da célula muscular cardíaca e permite a saída dos íons K+.
Desse modo, ocorre a redução da excitabilidade elétrica e dos batimentos cardíacos. A hidrólise do GTP a GDP pela subunidade α faz com que as subunidades se reassociem e a proteína G retorne ao estado inativo.

OBS: A proteína G também ativa enzimas que catalisam a síntese de moléculas sinalizadoras intracelulares (mensageiros pequenos ou segundos mensageiros). Uma dessas enzimas é a Adenilato-ciclase, responsável pela síntese do AMP cíclico (adenosina monofosfato cíclico) a partir do ATP (adenosina trifosfato). A enzima adenilato-ciclase é ativada pela subunidade α da proteína G. Essa enzima remove dois grupos fosfatos e reúne as extremidades da molécula (reação de ciclização). Desse modo, há um aumento na síntese do AMP cíclico em resposta à ativação da proteína G. Para eliminar o sinal, a enzima fosfodiesterase do AMP cíclico é ativada e converte o AMP cíclico em AMP.
1.2 -Receptores Associados a enzimas
Os receptores associados a enzimas atuam como enzimas ou associam-se a enzimas dentro da célula. Esses receptores foram descobertos devido à sua função em resposta aos fatores de crescimento, proteínas extracelulares que são responsáveis pela proliferação, diferenciação e sobrevivência celular. A maior classe desses receptores são os receptores tirosina-cinase (RTKs). Esses receptores possuem apenas um segmento transmembrana. Quando a molécula sinalizadora extracelular se liga, dois receptores se reúnem para formar um dímero na membrana. A função cinase é ativada pelo contato entre as duas caudas por fosforilação mútua das tirosinas presentes nas caudas dos receptores. Com isso, ocorre a agregação de um complexo de proteínas sinalizadoras intracelulares a essas tirosinas fosforiladas. Essas proteínas de sinalização intracelular tornam-se ativas e propagam o sinal.

Praticamente todos os receptores tirosina-cinase ativam a proteína Ras. Isso ocorre quando a proteína sinalizadora ligada à tirosinas fosforiladas recruta e estimula uma proteína ativadora de Ras. Essa proteína ativadora estimula Ras a trocar seu GDP por GTP. Ras ativada transmite, então, o sinal para diante na célula. Algum tempo depois, a ativa de GTPase de Ras causa a hidrólise de GTP a GDP e Ras retorna ao estado inativo.

Uma das vias estimuladas por Ras é a cascata de fosforilação da via das MAP cinases (Mitogenactivated protein kinase). Três proteínas-cinase conduzem o sinal para o núcleo da célula. No fim da cascata, a MAP cinase ativada fosforila proteínas efetoras que controlam a transcrição gênica e causam mudanças na atividade de algumas proteínas. Como resultado, ocorre proliferação celular, sobrevivência ou diferenciação celular.

Ras está mutada em grande parte dos tipos de câncer. A mutação inativa a ativade GTPásica de Ras e impede, portanto, que a proteína seja autoinativada. Com isso, acontece um aumento descontrolado da proliferação celular e desenvolvimento do câncer.
Outra importante via ativada pelos receptores tirosina-cinase é a via do Fosfoinositídeo-3-cinase (PI-3-cinase). A ligação do sinal de sobrevivência ao receptor tirosina-cinase, causa a ativação da enzima Fosfoinositídeo-3-cinase (PI-3-cinase). Essa enzima é responsável pela fosforilação do fosfolipídeo de inositol presente na face citosólica da membrana plasmática. Esses fosfolipídeos de inositol fosforilados, tornam-se sítios de ancoragem para proteínas sinalizadoras intracelulares, vindas do citosol para a membrana plasmática, que serão responsáveis por propagar o sinal.
Uma dessas proteínas sinalizadoras intracelulares que se liga ao fosfolipídeo de inositol fosforilado é a proteína-cinase B (PKB), também chamada Akt. Akt é uma serina/treonina-cinase que promove o crescimento e a sobrevivência celular. Ela atua, por exemplo, fosforilando e inativando a proteína citosólica pró-apoptótica Bad. Dessa forma, há inibição da apoptose e promoção da sobrevivência celular.

Ionotrópicos
Os receptores acoplados aos canais iônicos, também conhecidos como canais iônicos ativados de ligantes, medeiam direta e rapidamente a sinalização sináptica entre células eletricamente excitáveis (Fig. 3-4, A). Os neurotransmissores se ligam aos receptores e podem abrir ou fechar o canal iônico, dessa forma, mudando a permeabilidade iônica da membrana plasmática e alterando o potencial de membrana. Para exemplos e detalhes, consulte o Capítulo 2. Os GPCRs regulam a atividade de outras proteínas, como enzimas e canais iônicos (Fig. 3-4, B). Nesse tipo de receptor, a interação entre o receptor e a proteína alvo é mediada por proteínas G heterotriméricas, que são compostas por subunidades α, β, e γ. A estimulação das proteínas G, por receptores ligados a ligantes, ativa ou inibe proteínas alvos adiante na cadeia que regulam as vias de sinalização, se a proteína alvo for uma enzima, ou alteram a permeabilidade da membrana a íons, se a proteína alvo estiver em um canal iônico. Receptores catalíticos funcionam como enzimas ou estão associados a e regulam enzimas (Fig. 3-4, C). A maioria dos receptores acoplados a enzimas é uma proteinocinase ou está associada a proteinocinase, e a ligação ao ligante faz com que a cinase fosforile um subgrupo determinado de proteínas em aminoácidos específicos que, então, ativam ou inibem a atividade da proteína. Algumas proteínas da membrana não se enquadram na definição clássica de receptores; apesar disso, elas desempenham função semelhante à de um receptor, pois reconhecem sinais extracelulares e convertem esses sinais em segundo mensageiro intracelular que tenha efeito biológico. Por exemplo, quando ativadas por ligante, algumas proteínas de membrana passam por proteólise intramembrana regulada (PIR) que produz fragmento peptídico citosólico capaz de penetrar no núcleo e regular a expressão gênica. Nessa via de sinalização, a ligação do ligante a receptor de membrana plasmática leva à eliminação do ectodomínio, facilitada por membros da família metaloproteina.


b) Os hormônios que podem atravessar a membrana plasmática das células-alvo têm os seus receptores localizados no núcleo celular. Os hormônios devem atravessar a membrana plasmática e o citosol até chegar ao núcleo. A interação hormônio-receptor altera diretamente a transcrição de genes específicos, o que requer tempo para síntese de RNAm no núcleo e a subsequente síntese de proteínas nos ribossomos. Os hormônios são transportados ligados a proteínas específicas, e os esteroides e hormônios tireoideanos, utilizam este mecanismo e o tempo de ação é de horas e até dias.
B)
** **As moléculas de sinalização intracelulares são ativadas em resposta à ligação de uma molécula de sinalização celular em um receptor presente na superfície celular. As vias de sinalização intracelular são responsáveis por transmitir e propagar o sinal para dentro da célula; amplificar o sinal recebido e torná-lo mais forte; integrar sinais de diferentes vias de sinalização; e distribuir o sinal para mais de uma via de sinalização ou proteína efetora.
Na primeira classe, a proteína é ativada em resposta a uma proteína-cinase que adiciona um grupo fosfato à proteína e desativada em resposta à remoção do grupo fostato por uma proteína fosfatase.

Na segunda classe de proteínas, a ligação ao GTP ou GDP controla a atividade da proteína. Quando o GTP se liga, a proteína muda do estado inativo para o estado ativo. A atividade GTPásica da própria proteína faz com que ela seja inativada com a hidrólise do GTP a GDP.

A ação de hormônios, mediados por receptores nucleares, ocorre quando, após atravessarem a membrana plasmática por difusão simples, pois são constituídos de moléculas lipofílicas, entram no citosol e alcançam o núcleo. O complexo formado da união hormônio-receptor ativado, se une a regiões específicas do DNA para ativar ou inativar genes específicos, afetando a transcrição e a produção do mRNA respectivo. Existe um elemento sensível a hormônio (HRE) na região regulatória do DNA, perto do elemento promotor que regula a freqüência da iniciação da transcrição, de forma similar aos genes facilitadores (enhancers). O RNAm é depois traduzido nos ribossomos para produzir a proteína específica que causa a resposta metabólica.
As sequências de DNA dos HRE, aos quais se une o complexo hormônio receptor, são similares em comprimento, porém diferentes em sequência para os hormônios esteroidais. Para cada receptor, há uma sequência-consenso, à qual se une o complexo hormônio receptor. Cada sequência-consenso de HRE consiste de duas sequências de seis nucleotídeos, que podem estar vizinhas entre si ou separadas por três nucleotídeos. A habilidade de determinado gene em alterar a expressão de um gene em determinada célula depende da sequência exata de HRE e sua posição relativa no gene, bem como da quantidade de HREs associados ao gene.


Segundos Mensageiros
Os segundos mensageiros são substâncias que tem suas concentrações elevadas dentro das células em resposta a um hormônio primário, com a função de transmitir o sinal primário hormonal e traduzi-lo em alterações metabólicas dentro da célula-alvo. Como exemplos de segundos mensageiros, têm AMPc, GMPc, cálcio, proteínas-quinases, derivados do fosfatidil-inositol e a ação hormonal mediada por receptores nucleares.
AMPc:
O AMPc é o mediador comum da ação de muitos hormônios. É formado pela ativação de uma enzima plasmática, adenilciclase, que converte ATP em 3-5-adenosina monofosfato cíclica (AMPc), como consequência da interação entre um hormônio e seu receptor específico. Esta enzima pode ser estimulada ou inibida, mediante mecanismos que envolvem complexos protéicos regulatórios localizados na membrana plasmática ou proteína reguladora do nucleotídeo guanina, conhecidos como “Gs”, e “Gi”, que possuem subunidades α, β e γ. As “Gs” estão localizadas no lado citosólico da membrana plasmática, e se unem a um nucleotídeo, o GTP (trifosfato de guanosina), estimulando a produção de AMPc, pela ativação da adenilciclase. A proteína Gs está inativa, quando a subunidade está unida ao GDP. Quando ocorre a união hormônio-receptor, ocorre a fosforilação do GDP em GTP, tornando a Gs ativada. As subunidades, β e γ, dissociam-se da subunidade α. A subunidade Gs α, quando unida ao GTP, se desloca na membrana, desde o receptor até uma molécula de adenilciclase, ocorrendo sua ativação. Depois de ativada, a adenilciclase catalisa a produção de AMPc a partir de ATP.
Quando a subunidade Gsα se reassocia com as subunidades, β e γ, a Gs torna a estar disponível para uma nova interação com o complexo hormônio-receptor. O sinal continua dentro da célula com a união do AMPc a uma proteína quinase (proteína quinase A) que ativada, fosforila uma proteína, nos grupos hidroxila dos aminoácidos Thr e Ser, e esta proteína pode induzir mudanças em rotas metabólicas. A ação das proteínas-quinases é reversível pela ação de fosfatases específicas, as quais defosforilam as proteínas substrato das proteínas-quinases inativando-as. O estado de fosforilação ou defosforilação das proteínas substrato é o que determina sua atividade fisiológica. Um exemplo seria a enzima que degrada o glicogênio, a glicogênio-fosforilase a, que é ativa quando é fosforilada, enquanto que a enzima que sintetiza o glicogênio, a glicogênio-sintetase, é ativa quando defosforilada. Como as diferentes células têm receptores específicos para os diferentes hormônios, o AMPc opera como um metabólito comum para a ação de vários hormônios. Assim, cada célula tem diferentes enzimas que reconhecem diferentes hormônios, mas que são afetadas pelo AMPc.
Alguns hormônios atuam inibindo a adenilciclase, diminuindo os níveis de AMPc, evitando a fosforilação de proteínas específicas. Estes hormônios ao se unirem a seus receptores específicos ativam uma proteína G inibidora (Gi), homóloga estruturalmente a Gs. Ela atua de forma similar a Gs, se unindo ao GTP para ativar-se, porém ocorre a inibição da adenilciclase, diminuindo os níveis de AMPc. As catecolaminas e adrenérgicas, insulina, glucagon somatostatina e as prostaglândinas PGE1 e PG2, agentes opiáceos, LH, FSH, TSH, hCG, GnRH, PTH, calcitonina, utilizam este mecanismo.

Figura 3. Mecanismo de ação regulado pelo AMPc.
GMPc:
O guanosina-monofosfato cíclico (GMPc), é outro nucleotídeo que atua como segundo mensageiro, especialmente nas células do epitélio intestinal, coração, vasos sanguíneos, cérebro e dutos coletores renais. A ação GMPc, varia conforme o tecido. É derivado da conversão da guanosina trifosfato (GTP), pela ação da enzima guanilciclase. Os níveis de GMPc são 5% dos níveis de AMPc e podem ser aumentados pela ação de vários hormônios ou neurotransmissores, com acetilcolina, insulina, somatostatina, angiotensina e prostaglandinas, com isso acredita-se que o GMPc é intermediário de efeitos opostos aos efeitos do AMPc. Em muitos tecidos, os nucleotídeos cíclicos têm um papel contrastante, como na contração e relaxamento muscular. O GMPc está relacionado com a contração da musculatura lisa, e o relaxamento com a elevação do cAMP. Em tecidos excitáveis, como os músculos ou células nervosas, a despolarização é correlacionada com GMPc, e a hiperpolarização é relacionada ao AMPc. Existem substâncias que atuam através do GMPc. O fator natriurético atrial (ANF) é produzido por ativação da guanilciclase das membranas das células atriais do coração quando ocorre um aumento do volume circulatório de sangue, ocasionando uma dilatação do átrio. O ANF, também ativa a guanilciclase nas células coletoras dos túbulos renais, para aumentar a excreção de Na+, aumentando a excreção de água, diminuindo o volume circulatório. A pressão sanguínea é reduzida pela ação do ANF, mediante a GMPc, causando vasodilatação.
Nas células do intestino, um receptor de membrana que atua com a guanilciclase, pode ser ativado por uma toxina bacteriana, produzida pela E. coli, resultando em aumento da absorção de água na luz intestinal, ocasionando diarréia. A forma isoenzimática da guanilciclase no citosol é uma proteína associada ao grupo heme, estimulada pelo óxido nítrico, produzido a partir da arginina, pela ação da enzima NO-sintetase, dependente de Ca+. O GMPc produzido pela ação da guanilciclase, estimulada pelo óxido nítrico, causa diminuição da contração cardíaca. Ocorre um estímulo da bomba iônica que mantém baixa a concentração de Ca+ no citosol da célula cardíaca. Em muitos casos, o aumento dos níveis do GMPc é estimulado pelo fluxo de Ca+ no interior da célula, porque possivelmente esse íon é ativador da guanilciclase. O GMPc, assim como o AMPc, é hidrolisado porfosfodiesterases específicas.

Cálcio
O cálcio é um importante regulador de vários processos celulares, e atua também como um segundo mensageiro de ação hormonal. É essencial para ativação da fosfolipase A2, sendo requerido para a transdução do sinal entre o receptor hormonal e a adenilciclase ou a guanilciclase. Hormônios podem translocar Ca2+ para o citosol que funciona tanto nos processos de ativação enzimática, ou contração celular. Ele pode agir como um inibidor da atividade da adenilciclase, e pode aparecer como estimulador da atividade cíclica da fosfodiesterase dos nucleotídeos. O Ca2+ é requerido em alguns sistemas celulares, pois promove a interação entre o receptor hormonal e os nucleotídeos. A concentração de Ca2+ extracelular é maior que a intracelular (5 mM vs. 0,1-10 mM respectivamente). A concentração citosólica de Ca2+ é mantida em baixa concentração, mediante uma bomba de Ca2+ no retículo endoplasmático, nas mitocôndrias e nas membranas plasmáticas, sendo a entrada de Ca2+ é restrita e ocasionada por estímulos neuronais ou hormonais. A ação do Ca2+ é regulada por uma proteína chamada de calmodulina, ou proteína reguladora cálcio dependente (CDR). Ela possui quatro sítios de união com Ca2+, os quais provocam uma mudança conformacional quando estão ocupados, relacionada com a habilidade da calmodulina para ativar ou inativar enzimas.

Os co-fatores algumas vezes têm papel estrutural não-catalítico em certas enzimas, ligando juntas diferentes regiões da enzima para formar a estrutura terciária. Eles também podem servir como substratos que são quebrados durante a reação.
Derivados do fosfatidil-inositol
Na membrana plasmática, existe uma enzima hormônio-sensível chamada de fosfolipase C, que atua especificamente sobre o fosfotidil-inositol-4,5-difosfato, catalisando sua hidrólise em diacilglicerol (DAG) e inositol-1,4,5-trifosfato (ITP).
Esses dois compostos podem agir como segundo mensageiro da ação hormonal. O diacilglicerol é um ativador de proteínas-quinases, fosforilando proteínas específicas na membrana plasmática, e no citoplasma. O inositol trifosfato libera íons de cálcio armazenados
no retículo endoplasmático, com isso acredita-se que o ITP seja o integrador entre o hormônio e a mobilização de cálcio das reservas intracelulares. Estes mecanismos têm um papel importante no metabolismo do ácido araquidônico, precursor de algumas prostaglandinas. TRH, ACTH, LH, angiotensina II, serotonina e vasopressina, são alguns dos hormônios que atuam por este mecanismo.
Proteínas-quinases como intermediários da ação homonal
As proteínas-quinases são enzimas multifuncionais presentes na membrana plasmática. Um fator comum nos sinais de transdução hormonal seja através da adenilciclase, guanilciclase, cálcio/calmodulina, fosfolipase C, é a regulação sobre a atividade de uma proteína-quinase. É uma enzima multifuncional, presente na membrana plasmática das células. Existem centenas de proteínas-quinases, cada uma com seu sítio ativador específico e sua própria proteína substrato. A adição, de grupos fosfatos a resíduos dos aminoácidos Ser, Thr ou Tyr, introduz grupos carregados eletricamente em uma região moderadamente polar. Quando a modificação ocorre em uma região crítica para a estrutura tridimensional da proteína, devem ocorrer modificações em sua conformação, com isso, modificando sua atividade catalítica. Como resultado, os resíduos dos aminoácidos Ser, Thr ou Tyr, que podem ser fosforilados, e estão localizados em sequências-consenso da proteína, isto é, sequências repetidas que são reconhecidas pela proteína-quinase específica. Para poder servir como um mecanismo regulatório efetivo, a fosforilação causada pelas proteínas-quinases deve ser reversível, de modo a permitir o retorno ao nível anterior de estimulação quando o sinal hormonal terminar.
Segundos Mensageiros (2)
Derivados do fosfatidil-inositol
Na membrana plasmática, existe uma enzima hormônio-sensível chamada de fosfolipase C, que atua especificamente sobre o fosfotidil-inositol-4,5-difosfato, catalisando sua hidrólise em diacilglicerol (DAG) e inositol-1,4,5-trifosfato (ITP).
Esses dois compostos podem agir como segundo mensageiro da ação hormonal. O diacilglicerol é um ativador de proteínas-quinases, fosforilando proteínas específicas na membrana plasmática, e no citoplasma. O inositol trifosfato libera íons de cálcio armazenados no retículo endoplasmático, com isso acredita-se que o ITP seja o integrador entre o hormônio e a mobilização de cálcio das reservas intracelulares. Estes mecanismos têm um papel importante no metabolismo do ácido araquidônico, precursor de algumas prostaglandinas. TRH, ACTH, LH, angiotensina II, serotonina e vasopressina, são alguns dos hormônios que atuam por este mecanismo.
Proteínas-quinases como intermediários da ação homonal
As proteínas-quinases são enzimas multifuncionais presentes na membrana plasmática. Um fator comum nos sinais de transdução hormonal seja através da adenilciclase, guanilciclase, cálcio/calmodulina, fosfolipase C, é a regulação sobre a atividade de uma proteína-quinase. É uma enzima multifuncional, presente na membrana plasmática das células. Existem centenas de proteínas-quinases, cada uma com seu sítio ativador específico e sua própria proteína substrato. A adição, de grupos fosfatos a resíduos dos aminoácidos Ser, Thr ou Tyr, introduz grupos carregados eletricamente em uma região moderadamente polar. Quando a modificação ocorre em uma região crítica para a estrutura tridimensional da proteína, devem ocorrer modificações em sua conformação, com isso, modificando sua atividade catalítica. Como resultado, os resíduos dos aminoácidos Ser, Thr ou Tyr, que podem ser fosforilados, e estão localizados em sequências-consenso da proteína, isto é, sequências repetidas que são reconhecidas pela proteína-quinase específica. Para poder servir como um mecanismo regulatório efetivo, a fosforilação causada pelas proteínas-quinases deve ser reversível, de modo a permitir o retorno ao nível anterior de estimulação quando o sinal hormonal terminar.
Bibliografia
-
Transdução de sinais: uma revisão sobre proteína G Signal transduction: a review about G protein Priscila Randazzo de Moura1, Felipe Augusto Pinto Vidal2 1 Doutora em Farmacologia. Docente da Disciplina de Farmacologia, Instituto de Ciências da Saúde, Universidade Paulista (UNIP), Campus Sorocaba, SP. 2 Graduando do Curso de Farmácia, Instituto de Ciências da Saúde, Universidade Paulista (UNIP), Campus Sorocaba, SP.
-
https://www.ufrgs.br/lacvet/site/wp-content/uploads/2013/10/a%C3%A7ao_hormoniosTamara.pdf
-
https://www.scielo.br/scielo.php?script=sci_arttext&pid=S0004-27302001000300004
-
http://fisio2.icb.usp.br:4882/wp-content/uploads/2016/02/MECANISMO-DE-A%C3%87%C3%83O-HORMONAL.pdf